| Location: Home > Research > Research Progress |
| Out of Tibet: Ancestral Woolly Rhino Suggests Origin of Ice Age Megaherbivores in High Plateau |
|
Ice Age megafauna have long been known to be associated with global cooling during the Pleistocene and their adaptations to cold environments, such as large body size, long hair and snow-sweeping structures, are best exemplified by the woolly mammoths and woolly rhinos. These traits were assumed to have evolved as a response to the ice sheet expansion. In Science on September 2, 2011, Tao Deng from Institute of Vertebrate Paleontology and Paleoanthropology(IVPP), Chinese Academy of Sciences, and his colleagues from IVPP, Natural History Museum of Los Angeles County, University of Helsinki, Florida State University, University of Southern California, University of Texas, and Gansu Provincial Museum, report a new Pliocene mammal assemblage from a high altitude basin in western Himalaya, including a primitive woolly rhino. The new Tibetan fossils suggest that some megaherbivores first evolved in Tibet before the beginning of the Ice Age. The cold winters in high Tibet served as a habituation ground for the megaherbivores, which became pre-adapted for the Ice Age, successfully expanding to the Eurasian mammoth steppe.
Among the most iconic Ice Age mammals, the woolly rhino (Coelodonta) was widespread in northern Eurasia and adapted to cold climates in the mammoth steppe during the late Pleistocene. The known fossil record suggests that the woolly rhino evolved in Asia, but its early ancestry remains elusive. The new middle Pliocene (~3.7 million years ago) woolly rhino, Coelodonta thibetana, from the high-altitude Zanda Basin at the foothill of the Himalayas in southwestern Tibet occupies the most basal position of the Coelodonta lineage and is the earliest representative of the genus. As the Ice Age began about 2.8 million years ago, the Tibetan woolly rhino descended, through intermediate forms, to low altitude, high latitude regions in northern Eurasia, and along with the Tibetan yak, argali, and bharal, became part of the emerging Mammuthus-Coelodonta fauna in the middle to late Pleistocene. The holotype of the new species Coelodonta thibetana is a complete skull and mandible with associated atlas, axis, and third cervical vertebra, representing a full adult individual, discovered by Xiaoming Wang on August 22, 2007, and collected by a field team led by Gary Takeuchi.
The fossil locality is 10.2 km northeast of the Zanda county seat, Ngari District, Tibet Autonomous Region, China, with an elevation of 4,207 m. This holotype is from the upper part of the middle fine-grained sequence in the upper Zanda Formation, with an estimated age of 3.7 Ma in the middle Pliocene according to faunal and paleomagnetical dates.
Coelodonta thibetana possesses typical characteristics of the genus, such as a dolichocephalic skull, an ossified nasal septum, an enlarged, mediolaterally compressed nasal horn boss with a longitudinal crest, nasals bent downward anteriorly, occipital crest elevated and posteriorly extended, high tooth crowns with rough enamel and coronal cement, and visible medi- and post-fossettes at an early stage of wear. It differs from other, more derived species of woolly rhinos (C. nihowanensis, C. tologoijensis, and C. antiquitatis) in having a less well-developed nasal septum, which is only 1/3 of the length of nasal notch, a more anteriorly positioned mandibular symphysis, less derived molar morphology with thinner coronal cement, a less wavy ectoloph profile with a less prominent mesostyle on M2, a triangular occlusal outline of M3, a blunt anterior edge of the paraconid and a sigmoidal hypolophid with a distinctly recurved posterior end, and weak anterior ribs on m2-3.
The skull of Coelodonta thibetana is highly dolichocephalic (width/length ratio= 0.39), with a relatively long face as is characteristic of the genus. As in Coelodonta nihowanensis and C. antiquitatis, the nasal horn boss is strongly roughened and relatively large, occupying the entire dorsal surface of the nasal, indicating a large nasal horn. A wide and low dome on the frontals indicates the presence of a smaller frontal horn. The relative size of the nasal horn is greater than in extant and fossil rhinocerotines, resembling those of elasmotheres or dicerotines, but narrower in shape. As is typical of rhinoceroses with a low-slung head position for feeding at ground level, the occipital surface is reclined posteriorly, and the occipital crest is straight and backward, overhanging the occipital condyle to a similar extent as in C. nihowanensis and less than in more derived woolly rhinos.
The phylogenetic analysis of 5 extant and 13 extinct Rhinoceratini taxa places Coelodonta thibetana as a derived dicerorhine, nested within a clade of Mio-Pleistocene species of Stephanorhinus. The larger and heavier species such as Dihoplus ringstroemi, D. megarhinus and “D”. kirchbergensis consistently appear outside a clade including the more lightly built forms of Stephanorhinus (S. etruscus, S. hundsheimensis, S. hemitoechus) plus Coelodonta. In all of the most parsimonious trees the sister taxon of the Coelodonta clade is S. hemitoechus, a middle to late Pleistocene species. Within the Coelodonta clade, the phylogenetic analysis produced a series of progressively more derived species from C. thibetana, through C. nihowanensis and C. tologoijensis, and terminating in C. antiquitatis.
The carbon and oxygen isotopic compositions of fossil horse, rhino, deer, and bovid from Zanda Basin as well as a modern Tibetan wild ass (Equus kiang) were analyzed. The enamel-δ13C values for the time period of 3.1–4.0 Ma are -9.6+0.8‰ (n=110), indicating that these ancient herbivores, just like modern Tibetan asses in the area, primarily fed on C3 vegetation and lived in an environment dominated by C3 plants. The enamel-δ18O values of mid-Pliocene obligate drinkers (i.e., horse and rhino) are lower than those of their modern counterpart, likely indicating a shift in climate to much drier conditions after ~3-4 Ma. Using the δ18O values of fossil gastropods, the paleoelevation for the Mio-Pliocene Zanda Basin was estimated to be at least as high as, and possibly up to 1.5 km higher than today. This high elevation estimate is consistent with a cold (indistinguishable from modern) climate with lengthy subfreezing temperatures during the winters.
As in the woolly mammoth and the modern Tibetan yaks, the presence of long, woolly fur for thermal insulation in Coelodonta antiquitatis (a descendant of C. thibetana) strongly suggests that it was adapted to life in the cold tundra and steppe. In addition to using thick wool and increased body size to conserve heat, the cranial and horn complex in all woolly rhinos are also well integrated for cold conditions. While the classic notion that the long, forward-leaning and bilaterally flattened horn of the woolly rhino was used to “plough” through icy snow in order to feed during the winter is untenable for multiple reasons, the possibility that the horn was used to expose ground vegetation by sweeping loose snow is entirely plausible. Several morphological features are in favour of such an interpretation: 1, the horn of the woolly rhino tend to be more forwardly inclined (seen in preserved Ice Age cave paintings), usually to the point that the top portion of the horn is in front of the nose tip; 2, frequent presence of an anterior wear facet along the leading edge of the horn; 3, this facet is usually keeled, consisting of a right and left wear facet, presumably produced by a sweeping motion of the tilted head as the horn swept over snowy vegetation; 4, the bilateral flattening (i.e., anteroposterior elongation) of the horn, in contrast to the mostly rounded cross section in most living rhinos, gives the woolly rhino added profile for increased effective sweeping area; 5, posteriorly reclined skull (occiput) to assist in lowering of the head. These cranial features, in combination with long and woolly hair, are strong indications for a species adapted for survival in cold, snow-covered terrains.
The skull of the Zanda woolly rhino shows signs of the snow-sweeping morphologies. A large nasal horn boss indicates a differentially enlarged nasal horn relative to the frontal horn. This attachment area of the nasal horn shows a weak central “bump”, proximally and distally flanked by a weakly developed version of the central crest that characterises this anatomical region in the Pleistocene species. This arrangement indicates that the nasal horn was already bilaterally flattened at least to some degree. Furthermore, the downwardly curved nasal tip indicates a strongly forwardly inclined horn, and the posteriorly reclined occiput indicates a habitually low-slung cranial posture for feeding at ground level. Collectively, these cranial features lend support for snow clearing adaptations, which became a key pre-adaptation for its Pleistocene descendants.
The last representatives of the woolly rhino disappeared at the end of the Pleistocene, about 0.01 Ma. In addition to the new species from Tibet, only three species are commonly recognized: C. nihowanensis from the early Pleistocene (about 2.5 Ma) of northern China, C. tologoijensis from middle Pleistocene (about 0.75 Ma) of the Lake Baikal area of eastern Siberia to middle Pleistocene of western Europe, and C. antiquitatis from the late Pleistocene of northern Eurasia. All known species of the woolly rhino lived in cold conditions in northern Eurasia, especially Siberia, and a few rare southern forms are from high elevation regions, such as Aba, Gonghe, and Linxia in or along the eastern margin of the Tibetan Plateau. Such a striking zoogeographic pattern of progressive expansion from Tibet, coupled with the congruence of phylogeny and chronology, suggests a scenario in which ancestral woolly rhinos from high altitude Tibet descended to high latitude Siberia. As global climate cooled and cold habitats expanded, the ancestral woolly rhino descended to northern latitudes, and, eventually turning to grazing in the late Pleistocene, became one of the most successful Ice Age megaherbivores.
The woolly rhino is likely not the only case of a Tibetan cold-adapted ancestor to give rise to Ice Age megafauna. A composite Zanda fauna, as well as Mio-Pliocene records from elsewhere in the plateau, suggests that a distinct Tibetan fauna was in place going back as far as late Miocene. A progenitor of Tibetan bharal (Pseudois nayaur) appeared in the Zanda Basin and expanded to northern Asia during the Ice Age, a very similar history to that of the woolly rhino. Furthermore, molecular phylogenies for the Tibetan yak (Bos mutus) and central Asian argali (Ovis ammon) suggest links between presumed Tibetan (or surrounding mountains) ancestors and their North American megafauna relatives, such as the bison (Bison bison) and bighorn sheep (Ovis canadensis). Woolly rhino-like in its large size and woolly hair, the Tibetan yak has been recognized as far north as Lake Baikal area during the Pleistocene. Thus, while the origin of the cold-adapted Pleistocene megafauna has usually been sought either in the circumpolar tundra or in the cool steppes of the Pliocene and early Pleistocene, the harsh winters of the rising Tibetan Plateau could well have provided the initial step towards cold-adaptation for several subsequently successful members of the late Pleistocene mammoth fauna of the Holarctic.
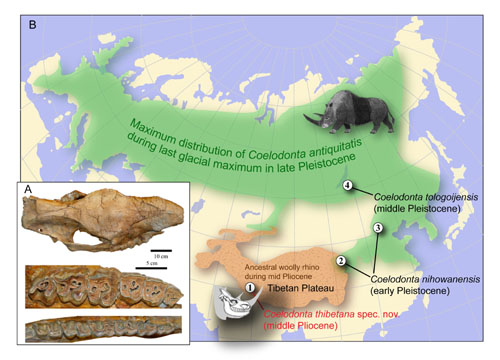 Fig. 1: A, Skull and cheek teeth of Coelodonta thibetana. B, Origin, distribution, and dispersal of woolly rhinos in Eurasia.(Image by DENG Tao)
Fig. 2: A, Phylogenetic position of Coelodonta thibetana. B and C, Enamel δ13C and δ18O values of herbivores in Zanda. D-F, three large mammals from Zanda fauna: D, Blue sheep; E, Snow leopard; F, Tibetan antelope. (Image by DENG Tao)
Fig. 3: Reconstruction of Coelodonta thibetana (Illustrated by Julie Naylor) |

